400-029-0925
400-029-0925胶质瘤干细胞和肿瘤微环境。GSCs的起源是有争议的,在不同的情况下可能会有不同的发生机制,但GSCs的形成高度依赖于TME及其旁分泌信号网络,无论是来自正常神经干细胞的突变还是来自体细胞GBM细胞的去分化。这些信号分子与肿瘤细胞的内部通道相互作用,促进肿瘤细胞基因组或表观组的不稳定,促进GSCs的形成和维持。
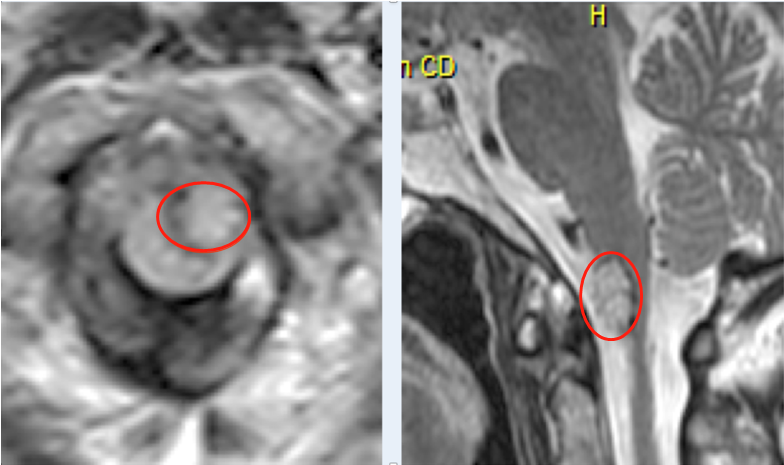
临床胶质瘤标本的免疫组织化学结果表明,GSCs只存在于胶质瘤的特定壁龛中,而不是均匀分布在整个肿瘤中。这表明,实体肿瘤的微环境条件对调节GSCs的分子和生物特性重要。因此,研究GSCs及其与微环境的交互作用不仅对理解GSCs的生物特性具有重要意义,而且可以评估GSCs对治疗的反应,并根据耐药肿瘤细胞确定新的治疗目标。
胶质瘤干细胞
1、GSCs和缺氧微环境
缺氧是肿瘤微环境的重要特征。GBM的快速生长使细胞处于相对缺氧的环境中。同时,低氧水平可以通过激活血管内皮生长因子来促进肿瘤细胞的生长。在胶质瘤中,VEGF可以诱导细胞增殖,促进细胞入侵,维持GSCs的不分化状态。
研究表明,缺氧可以促进单个分化的胶质瘤细胞形成克隆球,诱导干细胞标志物Sox2.Oct4.Nanog.CD133等表达增加。在低氧状态下,细胞会稳定地表达1α(hypoxia-induciblefactor1α、HIF1α)。发现HIF1α可以促进胶质瘤细胞在低氧条件下的分化。
Notch.STAT3等信号通路在维持GSCs的干性方面发挥着重要作用。HIF1α可以与NICD相互作用,使其稳定,从而激活Notch信号;缺氧可以激活STAT3信号通路,STAT3也是HIF1α的启动分子,诱导其RNA转录和蛋白质表达。
较近的研究表明,缺氧可以诱导FAT1基因的表达,FAT1可以提高HIF1α.VEGF的表达,促进Sox2.Oct4.Nestin等干性标志物的表达,调节上皮细胞间充质转化(epithelial-mesenchymaltransition、EMT)。
2、GSCs和酸性微环境
在大多数肿瘤中,实体瘤的微环境pH值低于正常组织,可能会随着实体瘤体积的增加而降低。酸性微环境往往被认为与缺氧有关,但Fukumura等。在体内胶质瘤异种移植的研究中发现,大脑肿瘤中在pH降低但氧含量正常的微环境,表明低pH可能是微环境中独自存在的特征。酸性微环境可以提高GSCs的Oct4.Nanog.Olig2基因表达,促进VEGF.IL-8等细胞因子的生成,进而通过旁分泌促进胶质瘤细胞的生长。
Hu等发现,在酸性微环境下,GSCs的自我更新能力、线粒体活性和ATP的产生得到了提高,从而促进和维持了GSCs的干性。研究还发现,活化的1.25二羟基维生素D3(1α,25(OH)2D3)可以控制Sox2.Oct4.Nestin等干性标志物的表达。在酸性微环境下,25羟基维生素D3-24-羟化酶(25-hydroxytaminD3-24-hydroxylase,CYP24A1)表达高,能催化1α、25(OH)2D3的快速降解。
3、GSCs和HMGB1
死亡的肿瘤细胞或应激性肿瘤细胞会释放损伤相关分子模式的分子(damage-associtedmolecules,DAMPs)到TME。如高迁移率族蛋白B1(highmobilitygroupbox1、HMGB1)、热休克蛋白70(heatshockprotein70、Hsp70)、三磷酸腺苷(ATP)、钙网蛋白(calreticulin、CRT)等。在这些DAMPs中,HMGB1通常在生理条件下定位于细胞核。当肿瘤细胞受到刺激或化疗和放疗的影响时,HMGB1会移位并释放到细胞外基质中。
细胞外释放的HMGB1可与GBM表面高亲和力受体结合,包括晚期糖基化较终产物受体(receptorforadvancedglycationd-products、RAGE)、toll样受体(tollikereceptor、TLR)-2.TLR-4和TLR-9。当HMGB1与受体结合时,激活关键信号通路和免疫反应,参与GBM生长、分化、运动和凋亡的调节。
Cheng等。发现细胞内HMGB1的高表达能促进GBM细胞的增殖和入侵;HMGB1高表达的患者与低生存曲线有关;GBM释放的HMGB1可以通过自分泌激活AKT和ERK信号通路,从而提高GBM的入侵能力。这些信息表明,抗HMGB1治疗可能是GBM的一种新的治疗方法。有趣的是,Zhao等人在CD133阳性人胶质瘤细胞系中使用慢病毒转染来表达HMGB1,发现控制了肿瘤细胞的增殖,增加了细胞的凋亡。此外,Chen等人在胶质瘤细胞系U87中加入人重组HMGB1蛋白,OCt4.Nanog干性标志物表达上升。
4、GSCs和细胞外囊泡
外泌体是脂质双分子层细胞的外囊泡,直径为50-110纳米(extracelulularvesicles,EVs)。几乎全部与胶质瘤相关的细胞都会释放外泌体,并存在于TME中。以外泌体为载体,GSCs可以与TME中的其他细胞进行通信和物质交换。外泌体可携带与肿瘤相关的mRNA.miRNA和蛋白质分子,从而调节GSCs的生长、入侵和耐药性。
Skog等研究发现,GBM外泌体中携带的RNA和蛋白质可以促进肿瘤的生长;此外,血液检测GBM来源的外泌物可以为胶质瘤患者提供诊断信息和辅助治疗决策。Figueroa等研究发现,与GBM相关的充质干细胞外泌物通过转移mir-1587增加了GSCs的致瘤性。
Bronisz等研究发现,GBM分泌的外泌物可以将miRNA-1传递给其他GBM细胞,从而改变GBM的入侵和增殖能力;此外,GBM外泌物还可以传递给基质细胞,促进血管内皮的形成。vanderVos等使用活体荧光显微镜技术直接观察到GBM分泌的外泌物将RNA转移到附近的小胶质细胞。这表明外泌物可能是GBM控制周围环境的一种手段。
Setti等研究发现,与正常组织相比,氯离子细胞内通道1(chloridintracelularchanel-1、CLIC1)在GBM中表达,在预后差的患者中表达较高;CLIC1可以促进GBM的增殖和自我更新能力。此外,CLIC1可以通过GBM外泌体分泌细胞。从CLIC1表达的GBM细胞中提取的外泌体是GBM体外增殖和体内肿瘤移植的强诱导剂。
综上所述,对TME的研究让我们对TME在GSCs增殖、入侵、自我更新等方面发挥了更深的作用,TME可以促进GSCs表型的维护。但目前对GSCs和TME关系的理解还不多方位。是TME的存在促进了GSCs的形成,还是GSCs产生了TME还不清楚。
- 文章标题:胶质瘤干细胞和肿瘤微环境
- 更新时间:2022-01-26 14:23:52