400-029-0925
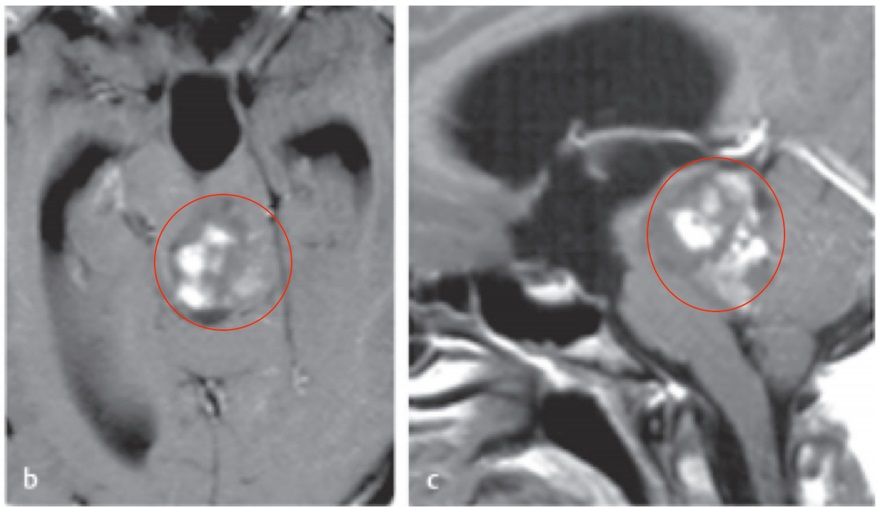
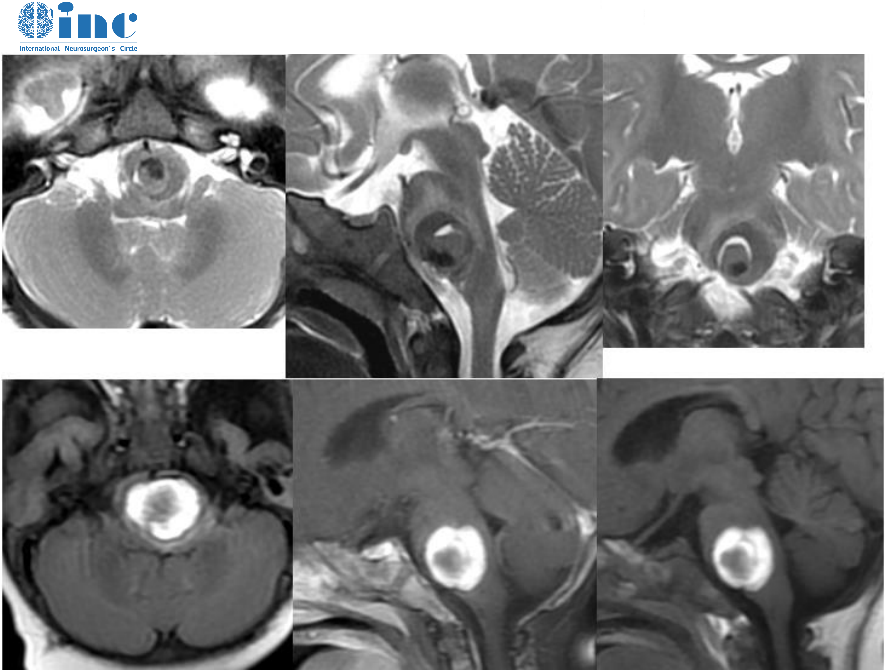
400-029-0925听神经瘤是一种常见的颅内肿瘤,大约占颅内肿瘤的8% -10%,主要起源于前庭神经鞘膜。由于前庭神经与面神经毗邻,肿瘤生长会对面神经造成牵拉、挤压损伤。
听神经瘤生长一般较缓慢,加之运动神经纤维可能对外来压力有较大的承受能力,所以手术前听神经瘤所致的周围性面瘫较少见。在手术中切除肿瘤时,肿瘤过大或进行面神经与肿瘤分离、电凝止血、神经牵扯等操作会加重对面神经的损; 常导致术后出现面瘫。而术后面瘫会对患者的生活、心理造成不良影响;因此面神经损伤的修复机制与治疗方法是目前神经外科研究的热点之一。现对听神经瘤切除术后的面神经损伤修复的研究进展综述如下。

1 面神经的相关解剖
面神经主要包括管理表情肌运动的运动纤维和中间神经。面神经核位于被盖的腹外侧,面神经核发出面神经的主要部分,在脑桥下边界与中间神经一同出脑干,支配面部表情肌。中间神经由两组副交感神经和两组不同感觉纤维组成;其中副交感神经根起源于上泌延核,其后为岩浅大神经和鼓索的组成成分,分别剌激泪腺、鼻咽颗部腺体与领下、舌下腺体的分泌。其两组的感觉神经根起自孤束核,其中的一组为味觉纤维通过鼓索,司舌前 2/3 的味觉。另一组为一般感觉纤维接受鼓膜、内耳、外耳道及周围皮肤的感觉。面神经在桥延沟外侧出脑干,与前庭蜗神经伴行,经过脑桥小脑池穿过蛛网模下腔到达内耳道。前庭蜗神经在耳蜗和骨迷路之间走行;蜗神经进入了耳蜗;面神经移行为膝状神经节并发出岩浅大神经后经鼓室段向乳突走行,较后经茎突孔出颅,之后走行于腮腺实质内,并分为颞支、颧支、颊分支、下颌支、颈支,进而支配相应的面部表情肌。在面神经与前庭蜗神经进入内耳道的相应神经管之前,内耳道的横崎将上方的前庭上神经和面神经与下方的前庭下神经和蜗神经分开,垂直嵴将前庭上神经与面神经分开,面神经位于前庭蜗神经的前上方。

面神经,图自网络
2 组织学
神经系统包括中枢神经系统和周围神经系统,周围神经系统义包括脑神经和脊神经;面神经属于周围神经系统。一些研究观察发现,面神经出脑桥后的数毫米的髓稍由少突胶质细胞包裹形成,之后经过一个中枢神经到周围神经系统( CNS-PNS) 过渡区,较后延续成施旺细胞包绕构成髓鞘的外周段。面神经由神经外膜包绕,神经外膜为一疏松的血管结缔组织鞘,包含有血管、淋巴管和多种神经束膜,并且包绕着许多神经纤维束并形成神经。每一神经纤维束又由神经束膜所包绕,施旺细胞再由网状胶原纤维形成的神经内膜所包绕。神经纤维分为有髓鞘和无髓销两种。在有髓鞘神经纤维中,施旺细胞为沿着轴突链状排列的神经纤维膜细胞,每 一个施旺细胞完全包裹轴突,细胞质膜会形成一个轴系膜,轴系膜不断的移动并且在轴突周围同心旋转包绕形成连续多层的细胞质膜,这些细胞膜较终形成髓销;施旺细胞之间不完全连接,此处较狭窄且使轴膜裸露,称为郎飞结。面神经的有髓和无髓鞘神经轴突都被施旺细胞包绕,施旺细胞外围还被基底膜覆盖包绕,基底膜外又面向神经内膜结缔组织。有髓鞘神经纤维中,每个施旺细胞在郎飞结的节间形成髓销,并沿着轴突间断直线排列;而且基底膜在施旺细胞甚至郎飞结的部位都是连续不断的 。当临近郎飞结时,髓鞘逐渐减少并变成类似于锥形的形态直至消失,较后只有细胞质膜形成的副结袋。而无髓鞘神经纤维的轴突则嵌入施旺细胞内,轴突周围为细胞质膜单层包绕,并形成系膜样的结构。神经的轴突内则包含微管、神经纤维细丝、细长光滑内质网、线粒体等结构。有髓鞘神经纤维是一根轴突,并由将轴突纳入施旺细胞鞘中的施旺细胞组成,无髓鞘神经纤维则是多个轴突被一个施旺细胞包裹。每一根神经纤维被基底层包裹形成神经内膜,以便于在结构上与其他神经纤维隔离开。轴突周围的分层的纤维骨架结构使神经更加稳定并且在延长使更加有弹性,还能更好地传播动作电位。
3 周围神经损伤后的再生过程
当周围神经受到挤压或者切割伤时,神经损伤的去神经断端远端发生沃勒变性( Wallerian degeneration)。神经损伤后施旺细胞表达与钙激动相关的磷脂酶,使部位远端的轴突和髓鞘溶解成无数个类似于"椭圆体"的碎片。沃勒变性时神经会释放一些可扩散的因子,导致神经纤维周围的毛细血管中的单核细胞进入损伤的神经内膜销中变成巨噬细胞,这些退化分解的髓鞘都被日噬细胞吞噬,巨噬细胞也可以触发施旺细胞增殖,并分泌许多细胞因子如 IL-l 可促进轴突生长 。神经损伤后,孤立的施旺细胞会从髓销形成模式变成增殖模式,并且对促细胞分裂素产生应。还有研究表明,在周围神经损伤后,一些施旺细胞会转变成修复形态的细胞占据神经损伤的远端,这些修复细胞专门支持轴突再生:所以处于修复形态的施旺细胞的持续存在,则成为神经再生成功的一个关键环节。较近有小鼠实验发现,转录因子 STAT3与修复施旺细胞的长期存在的维持有相关性。神经损伤引发施旺细胞中的 STAT3的激活,并支持长期去神经化的施旺细胞的生存和形态学的分化状态。这些处于修复状态的施旺细胞会在基底膜管道之中形成一个层状型组织的细胞链,被称作施旺细胞柱(即 bands of Büngner) ;施旺细胞柱的细胞质膜上会表达粘附因 (adhesion molecules) ,为轴突再生提供支架,还能为轴突再生提供营养因子。并且多个实验表明,施旺细胞的基底膜象施旺细胞柱一样也提供合适的支架;表明基底膜内表面也含有某些粘附分子,可与轴突合适的粘连;另外基底膜包绕所形成的管道为再生的轴突提供了一个合适的空间,以隧道的形式使之通过并生长。
神经受到横断切割伤的数小时内,在断端的近端轴突表面轴突会出芽式地产生许多小分支,这些小芽体常在损伤神经近端的郎飞结产生。并且上述芽体会逐渐发展成结构良好的生长锥(grocones),并向远端延伸;生长锥会被触角样的丝状伪足跨越,丝状伪足上的表面受体会与施旺细胞表面和施旺细胞基底膜上的表面秸附分子暂时地锚定,丝状伪足中的肌动蛋白丝附着于表面受体,以此予以生长锥产生向前的动力。因此在神经损伤远端的施旺细胞为生长锥的延伸提供导向,使生长锥贴近施旺细胞,并沿着其表面通过基底膜和施旺细胞之间的空隙延伸,较终进入神经远端部分。髓鞘也沿着近端再生的轴突开始生成;施旺细胞从近端迁移出来,并以小组为单位一对一的包绕轴突,在每一根轴突上开始形成髓销;等到神经再生完毕,郎飞结之间的髓稍长度会比损伤前的短。
4 影晌周围神经再生的因素
4.1 粘附分子在通过施旺细胞柱的延伸过程中,再生轴突的生长锥可以选择性地在质膜上表达出不同的粘附因子。轴突和施旺细胞之间的粘附可被免疫球蛋白超家族和钙粘蛋白的超家族等粘附因子调节。然而,轴突和基底膜之间是由层粘连蛋白调节。
4. 1. 1 免疫球蛋白超家族 (immunoglobulin superfamily) 轴突和施旺细胞的连接可部分被免疫球蛋白超家族调节,如神经细胞蒙古附分子 (neural cell adhesion molecule , N-CAM) LI 。在伴随着周围神经损伤去神经支配的时候, N-CAM LI 在施旺细胞表面重新表达,并且使施旺细胞相互连接形成施旺细胞柱,当重生的轴突长入施旺细胞柱, NcCAM LI也会在质膜上表达;并且作为轴突和施旺细胞之间连接的粘附分子以促进轴突生长。
4.1.2 钙粘蛋白超家族(cadherin superfamily) 钙粘蛋白 (cadherin) 是一种 Ca依赖的选择性细胞间粘附因子,通过与特定的细胞绑定,在神经再生中发挥重要作用。钙粘蛋白有很多子类,如在周围神经再生时,神经钙蛋白( N-cadherin) 在轴突和施旺细胞质膜上出现免疫反应;相反,在轴突质膜与基底膜上就没有这样的免疫反应;由此可见 N-cadherin节轴突与施旺细胞之间的粘附。
4. 1. 3 层粘连蛋白(laminin) 及其受体施旺细胞的基底膜属于细胞外基质,再生轴突优先粘附到基底膜,层粘连蛋白在基底膜中大量存在,是促进轴突生长较强有力的粘附分。实验表明,周围神经中层粘连蛋白的损害会导致施旺细胞的异常分化,有层粘连蛋白缺陷的施旺细胞导致其一直处于髓鞘形成前的阶段。所以层粘连蛋白对施旺细胞的去分化、增殖及髓销再生也至关重要。整合蛋白( integrin) 属于异质二聚体跨膜糖蛋白的大家族,并且由特定配对的。亚基(蛋白链)组成。层粘连蛋白和整合蛋白的结合绑定,可触发磷脂瞰肌醇-激酶 (PI 3-kinase) 的磷酸化;并进一步激活 AKT ,然后使糖原合成酶激酶布(glycogen synthase kinase-3? , GSK-3?) 磷酸化并导致 GSK-3?活性受控制;较终促使细胞骨架的重新排列和延伸,引起自发的和神经生长因子(nerve growth factor, NGF) 或胶质细胞源性神经营养因子 (glial cell line-derived neurotrophic factor, GDNF) 所诱导的轴突的再生生长。
4.2 神经营养因子( neurotroph ic factors) 神经营养因子对于调节施旺细胞分化和轴突髓鞘的形成起到至关重要的作用。周围神经损伤后的沃勒变性中去神经化的施旺细胞可产生神经营养因子来维持细胞存活,剌激轴突生长。这些营养因子包括 NGF ,脑源性神经营养因子( brain-derived neurotrophic factor, BDNF) 、神经营养素( neuro-trophin , NT) NT415 [11 剧。神经营养因子的信号通路是通过两种类型的受体介导,包括 Trk 受体和 p75NTR。 p75NTR 可以与NGF BDNF N1习、 NT-4/5 以相同的方式结合,然而 Trk体有 种不同的类型,每一个都专门与一种神经营养因子结合, TrkA 选择性地与NGF 结合,TrkB专门与 BDNF NT-41结合,TrkC优先与T3结合。目前对神经营养因子的研究基本处于基础实验阶段,其临床应用的效果还需要进一步的研究与论证。
4.2. 1 NGF NGF 是由交感和感觉神经的目标器官所合成和分泌的,并被神经末端通过受体介导胞吞作用摄取,通过轴突逆行运输到神经元,促进神经元的存活和分化。实验表明,在周围神经损伤后,远端施旺细胞和浸润到神经损伤部位的巨噬细胞都会产生 NGF。并且NGF已被证实可以保存施旺细胞,并借此促进损伤近端的轴突再生,调节轴突的生长和分叉NGF和其相应受体TrkA 的结合使细胞浆上的受体的酶氨酸残基磷酸化;随后,触发细胞内的信号级联反应,包括 MAPKl ERK PI-3 K1Akt phospholipase C ( PLC)--y 信号通路,以促进神经元的存活。 NGF TrkA 体结合后还可以激活 GSK-.布,使得 MAPIB 被磷酸化,从而调节生长轴突的微管动力。
4.2.2 BDNF NT415 实验表明,神经切断后4周时,其BDNF mRNA 的浓度比正常水平增高数倍。 BDNF 不仅促进感觉和交感神经的存活、生长,也促进运动神经元存活[l1 BDNF NT4/5 通过与其相应的受体绑定,通过 PI-3 K1 Akt MAPKl ERK 通路来激活 cAMP 应答元件结合蛋白 (cAMP response element binding protein , CREB) ,并且 BDNF NT4/还可以增加生长相关蛋白 43 ( growth associated protein43 , GAP43) 的表达, CREB GAP43 也都参与促进轴突再生的过程。
4.2.3 纤维母细胞生长因子 (fibroblast growth factor, FGF) 神经损伤后 FGF-2 被正调节并高表达。FGF-2 被敲除小鼠的研究表明,神经损伤时FGF-2 系统有助于施旺细胞的增殖和再分化的调节;在体外FGF-2 是施旺细胞的一种潜在促进分裂的分裂素。施旺细胞产生碱性 FGF( bFGF) , bFGF并与在基底膜上的蛋白聚糖( proteoglycans) 结合,可使bFGF在基底膜上压缩,并使 bFGF 容易促进与基底膜接触的再生的轴突生长。
4.3 生长锥的影响因素
生长锥一直作为负责轴突生长、导向、和目标的识别的不同结构。轴突的延伸是通过膜的成分供应到生长锥质膜而实施的,而不是表面轴膜的近端向远端流动。组织有序的细胞骨架结构与细胞内信号传导的联系决定生长锥的形状和能动性;细胞内钙离子调节生长锥的能动性。在轴突延长过程中,生长锥的生长活动和导向大部分被生长锥所粘附的基底物管理;生长锥通过利用蛋白酶溶解进行延伸,以通过结缔组织。引导信号分子通过与生长锥表面的受体结合发挥重要作用,触发二级信号并且引导轴突向正确的方向扩展。这些导向因子 (guidance cues) 括信号素( semaphorins )及其受体、神经纤毛蛋白( neuropilins) 等。 GAP43 是一个钙调节素结合蛋白,属于细胞骨架接头 (cytoskeletal adaptors) 的一种, GAP-43氨基末端与质膜相连,叛基端与细胞骨架成分相连。
GAP-4 可能是生长锥中的细胞内信号传导过程中可以被磷酸激酶调节的关键蛋白,以促进轴突再生。实验研究表明, GAP-4 可能与轴突的直接生长无关,而与生长锥的铺展、分叉、粘附有关。
5 面神经损伤的治疗
5.1 药物治疗
5. 1. 1 美金刚( Memantine) 美金刚一种不同的非竞争性NMDA受体拮抗剂,是金刚皖的一种衍生物,并且还是一种钙通道阻捕剂。在神经挤压伤后的神经缺血期间,由于细胞外谷氨酸诱导细胞内钙沉积和细胞凋亡,导致 NMDA 受体的激活。美金刚可以绑定到NMDA受体,并控制 NMDA体上谷氨酸的活性;从而控制细胞凋亡,促进面神经功能恢复;而且美金刚对其他非 NMDA受体没有任何影响。
5. 1. 2 尼莫地平( Nimodipine)
尼莫地平能够轻易地通过血-脑屏障,并且通过 型电压依赖的钙通道来对抗钙内流。 这个钙离子通道存在于脑干中的面神经核,并主要存在于树突。有实验表明,在面神经运动核的树突和面神经轴突可能有尼莫地平作用的靶点在活体轴突延长的实验表明,生长锥中钙爆发(钙爆发又称之为钙脉冲,来源于较低水平的剌激导致单个通道的短暂开放而释放出来的钙的频率的减少会增加神经再生的速度,所以尼莫地平可能通过调节钙离子流入到生长锥,而为神经出芽生长提供更好的环境,从而增加轴突和髓鞘的数量和厚度以促进轴突生长;其临床应用还有待进一步评估。
5. 1. 3 硫酸服基丁胶 (Agmatine sulphate) Surmelioglu将小鼠面神经下顿支横断之后,再将断端缝合;并给小鼠腹腔内注射硫酸肌基丁胶。通过对照实验和显微镜形态汁量学评估后表明,硫酸肌基丁胶联合神经缝合治疗与单纯的缝合治疗相比,前者可以更合适地促进神经再生。但是在临床治疗上如何应用硫酸肌基丁胶还需要进一步的研究。
5.2 手术治疗
5.2.1 舌下神经侧端与面神经末端吻合术及光生物调节疗 (PBMT) 对出现医源性面瘫的患者,将面神经纤维近端横断,并且在舌下神经的背侧面沿着斜后的方向切出一个切口,将面神经横断面远端与舌下神经背侧切口无张力吻合;并在缝合位点予以一层纤维蛋白胶覆盖。高颈位水平的舌下神经的背侧面含有-颈神经的交通支运动纤维。因此在舌下神经背侧面的切口可以在程度上避开负责语言功能的纤维,从而避免造成长期的语言功能障碍。但近期小鼠实验观察又表明,侧端舌下-面神经吻合后从神经形态学观察发现,虽然神经侧端存在出芽生长,但是神经末端出芽的生长才是神经再生纤维的主要来源;侧端舌下,面神经吻合后的神经形态学和功能的恢复的效果可能不如端端吻合。在神经缝合的位点还可以用光生物调节疗法治疗。光生物调节疗法治疗是利用嫁铝碑化合物激光以持续的脉冲作用于缝合位点,激光可以渗透进入神经组织,可以促进线粒悍的产生、增加血供、促进施旺细胞增殖和胶原纤维合成,并且还有抗炎、抗水肿的作用,从而促进神经再生。
5.2.2 咬肌-面神经吻合术 有研究报道,耳大神经是用于神经嫁接的较好的供体。因为耳大神经的缺损所导致的耳垂感觉缺陷的发病率较低。米兰大学加莱亚齐医院的Biglioli 等对肿瘤切除术后发生面瘫的7例患者行咬肌,面神经吻合术,将面神经出颈突孔的主干切断,并将咬肌神经远端横断,咬肌神经断端的近端与面神经断端的远端旋转对接;对接前将耳大神经嫁接到二者之间;对接点予以缝合,并用纤维蛋白胶覆盖包裹;术后患者均恢复了面部表情模仿的功能和面部两侧的对称性,恢复时间为2-9个月,平均为4.8个月;7例患者眼良险闭合时的巩膜显露从术前的平均5.7 mm恢复到术后的0. 7 mm 经麻痹患者(术前 HB 分级为 级)行咬肌-面神经吻合术,并将耳大神经或周围神经嫁接其中;经过术后个月的随访,5.9%的患者恢复到 HB1级,1.8%的患者恢复到到当习HB II级,20.6% 的患者恢复到 HBⅢ级;并且全部患者均未出现咬肌神经所涉及到的功能异常;但许多患者诉有耳垂或足跺小范围的感觉缺失。要使咬肌-面神经吻合术成为成熟、合适、顺利的临床治疗手段,仍有待进一步研究及反复临床试验论证。
6 总结与展望
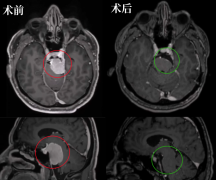
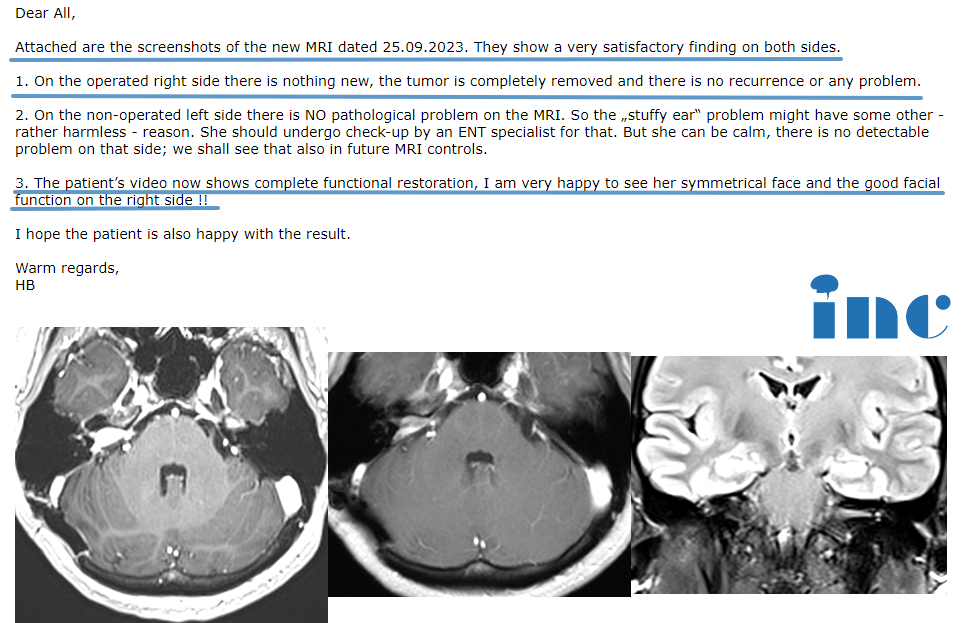
目前听神经瘤术后并发症仍是神经外科医生需要克服的难题,是对于听神经瘤术后面神经功能的保护和修复问题方面。对于面神经的保护,在听神经瘤切除术中至少要确定面神经的完整性,做到解剖保留。避免对面神经产生重度的牵拉,并避免电凝面神经。在面神经的修复方面,目前有营养神经药物和手术修复治疗。手术治疗如舌下-面神经吻合术、咬肌-面神经吻合术,这些术式对面神经的修复都有明显的促进作用,其效果在临床上已得到证明。在促进面神经修复的药物治疗方面,对神经修复过程中各个环节的分子机制的研究显得尤为重要;特别是对于信号通路及其对应的药物作用的靶点方面的研究,可能是面神经修复治疗研究方面的方向。
资料来源:DOI: 10. 3969/j. issn. 1672-7770.2019.01. 021
- 文章标题:听神经瘤术后面神经损伤修复的研究进展
- 更新时间:2021-07-01 11:38:02